 In the most recent 2015 issue of Christian Spirituality and Science published by Avondale College (a Seventh-day Adventist institution under the South Pacific Division of Seventh-day Adventists) there are three articles, all of which appear to challenge the firm position of the Seventh-day Adventist Church on the literal nature of the creation week described in Genesis. The authors of these papers suggest that the church should put aside such antiquated 19th century notions and accept or at least tolerate the idea that life has existed and evolved on this planet over the course of at least a couple billion years or so. Of course, this comes at a time when the church as an organization is considering strengthening its statement of “fundamental beliefs” by adding language to the existing statement on creation that makes the church’s understanding of the literal and recent nature the “creation week” quite clear and unambiguous.
In the most recent 2015 issue of Christian Spirituality and Science published by Avondale College (a Seventh-day Adventist institution under the South Pacific Division of Seventh-day Adventists) there are three articles, all of which appear to challenge the firm position of the Seventh-day Adventist Church on the literal nature of the creation week described in Genesis. The authors of these papers suggest that the church should put aside such antiquated 19th century notions and accept or at least tolerate the idea that life has existed and evolved on this planet over the course of at least a couple billion years or so. Of course, this comes at a time when the church as an organization is considering strengthening its statement of “fundamental beliefs” by adding language to the existing statement on creation that makes the church’s understanding of the literal and recent nature the “creation week” quite clear and unambiguous.
 Consider, for example, the article written by Dr. Lynden J. Rogers (Dean of the Faculty of Science and Mathematics at Avondale College) entitled, “Old Universe But Young Life?” In this paper professor Rogers briefly reviews the history of the church’s understanding of the origin of the Earth and the Universe. He then points out that many in the church leadership currently accept that the universe and even the material of the Earth is quite likely very old indeed! – while still holding to the notion that life on Earth must still be of recent origin. Rogers argues that these views are inconsistent with each other and are based on an inconsistent use of science that is simply irrational. To illustrate his point he cites several examples of why mainstream scientists believe the Earth, and life on it, must be very old – on the order of around 4.5 billion and 2 billion years respectively:
Consider, for example, the article written by Dr. Lynden J. Rogers (Dean of the Faculty of Science and Mathematics at Avondale College) entitled, “Old Universe But Young Life?” In this paper professor Rogers briefly reviews the history of the church’s understanding of the origin of the Earth and the Universe. He then points out that many in the church leadership currently accept that the universe and even the material of the Earth is quite likely very old indeed! – while still holding to the notion that life on Earth must still be of recent origin. Rogers argues that these views are inconsistent with each other and are based on an inconsistent use of science that is simply irrational. To illustrate his point he cites several examples of why mainstream scientists believe the Earth, and life on it, must be very old – on the order of around 4.5 billion and 2 billion years respectively:
 Theory of Plate tectonics: Supported by matching puzzle-fitting continents with actively spreading mid-oceanic ridges that must have taken place over vast periods of time when life was clearly in existence on the planet.
Theory of Plate tectonics: Supported by matching puzzle-fitting continents with actively spreading mid-oceanic ridges that must have taken place over vast periods of time when life was clearly in existence on the planet. Geologic Column: With its sequentially-deposited layers that must carry temporal implications which can’t be tenably explained by the usual creationist appeals to various sorting mechanisms such as hydrological sorting, ecological zonation, and relative mobility.
Geologic Column: With its sequentially-deposited layers that must carry temporal implications which can’t be tenably explained by the usual creationist appeals to various sorting mechanisms such as hydrological sorting, ecological zonation, and relative mobility. Radiometric Dating: With its ability to date igneous rocks and the fossils associated with those rocks with a “high degree of data concordance” is “compelling for most scientists” and is only supported by additional methods such as radiocarbon dating and amino acid racemization dating techniques. The usual creationist arguments which appeal to a significant reduction in the half-lives of the various radioactive isotopes is simply untenable as a viable rescue for the young-life perspective on origins.
Radiometric Dating: With its ability to date igneous rocks and the fossils associated with those rocks with a “high degree of data concordance” is “compelling for most scientists” and is only supported by additional methods such as radiocarbon dating and amino acid racemization dating techniques. The usual creationist arguments which appeal to a significant reduction in the half-lives of the various radioactive isotopes is simply untenable as a viable rescue for the young-life perspective on origins. Ice Core Dating: With the fairly recent ice-core studies of the vast ice sheets in Greenland and Antarctica showing extensive data regarding ancient volcanic activity and climate changes over hundreds of thousands of years – and backed up by very similar findings based on lake-bed sediment cores. Clearly life has been around this entire time since pollen grains and diatoms are spread from top to bottom within the ice cores and lake bed cores.
Ice Core Dating: With the fairly recent ice-core studies of the vast ice sheets in Greenland and Antarctica showing extensive data regarding ancient volcanic activity and climate changes over hundreds of thousands of years – and backed up by very similar findings based on lake-bed sediment cores. Clearly life has been around this entire time since pollen grains and diatoms are spread from top to bottom within the ice cores and lake bed cores.



And, these are only a handful of the examples that could be provided! Clearly then, the desire of the church to continue to hold to the concept of a literal interpretation of the first few chapters of Genesis is very much outdated and not at all scientifically tenable or otherwise rational – according to a professor teaching and heading the science department in one of the church’s own schools! One must either accept that life on Earth is old, or if one still believes that life was in fact created within just one literal week, to be consistent, one must therefore argue that the entire universe is young and was also created within that same week of creation. Of course, Professor Rodgers goes on to argue for God being the Designer of the universe, citing some of the numerous fine-tuned features of the universe necessary to support life. He also notes that several prominent physicists also argue favorably for the “anthropic” view of the universe with some kind of high-level intelligence behind it.
Rogers goes on to explain that there is a real problem with allowing for a “process-driven model for the universe”, starting with a “big bang”, while denying a similar process-driven model for the “biosphere” – i.e., for life and its evolution over time. He goes on to quote himself in an earlier article that he wrote for Spectrum:
. . ., cosmologists understand that the Big Bang singularity was just the beginning of a long process, which by all appearances was hugely violent, random and wasteful, but through which God has obviously produced a highly specific outcome – our anthropic universe. How do we feel about God implementing his grand design in such a protracted manner? Further, if as creationists we accept this view, do we in any way weaken the basis on which we might counter others who suspect that God may have used similar processes to develop life?

In other words, what’s wrong with neo-Darwinism and all the pain, suffering, and death, and general waste of life and resources required for Darwinian evolution to work if one already believes that God used essentially the same wasteful mechanism in the creation of the universe? Or to quote Rogers directly:
If God could set off the blue touch powder of the Big Bang in order to initialise this universe and set it on an inexorable path to a finely tuned state of suitability as a womb of life, why could not some similar event under His direction have initiated life itself and established its path to sentience and God-recognition? If it is theologically acceptable for God to use a developmental process for one, why not for the other? Conversely, if it is theologically unacceptable to entertain the idea of such a developmental sequence for life, then why is it acceptable for the universe?
 Rogers goes on to argue that living things on this planet likely evolved in a very similar manner to how the universe “evolves” – citing the theory that the first eukaryotic cell (cells that contain a nucleus) evolved through symbiotic means when prokaryotic bacteria were engulfed, but not digested, by the earliest ancestor to all eukaryotes. They eventually formed a symbiotic relationship and worked together to produce the ancestor of all eukaryotic creatures living today – giving rise to such things as mitochondria and chloroplasts.
Rogers goes on to argue that living things on this planet likely evolved in a very similar manner to how the universe “evolves” – citing the theory that the first eukaryotic cell (cells that contain a nucleus) evolved through symbiotic means when prokaryotic bacteria were engulfed, but not digested, by the earliest ancestor to all eukaryotes. They eventually formed a symbiotic relationship and worked together to produce the ancestor of all eukaryotic creatures living today – giving rise to such things as mitochondria and chloroplasts.
This fortunate invasion is understood to have enabled a “complexity of structure and function that neither component could have evolved separately.”
So, what does this all mean? Well, according to Rogers, what this means is that “God’s causality is then indistinguishable from natural law…” In other words, there are no such things as “miracles” in nature that cannot be explained by what we term “the laws of nature” or that would require a special act of “intelligent design” of any kind, much less an act of Divine creative power outside of the natural laws that God originally created, Rogers explains:
As scientists we can study the arrow in flight but science itself can show us neither the archer directing the process nor the end point – the target.
To read the full article: Link
__________
My take on Dr. Rogers’ article:
 My first thought after reading the article was that Mrs. White would be rolling over in her grave if she knew what the school she helped to found in the late 1800s was promoting in its “Christian Spirituality and Science” journal. How can one of our own schools be publishing articles like this that strike as the very basis of the church’s existence? – the very basis of the name “Seventh-day Adventist”? And, how can the Adventist Church continue to be associated with a school that is so willing to publicly thumb its nose that the organized church and its leadership?
My first thought after reading the article was that Mrs. White would be rolling over in her grave if she knew what the school she helped to found in the late 1800s was promoting in its “Christian Spirituality and Science” journal. How can one of our own schools be publishing articles like this that strike as the very basis of the church’s existence? – the very basis of the name “Seventh-day Adventist”? And, how can the Adventist Church continue to be associated with a school that is so willing to publicly thumb its nose that the organized church and its leadership?
 Beyond this, however, how good are the arguments presented by Dr. Rogers? Perhaps the church is wrong after all? Perhaps I should leave the SDA Church and Christianity as well? Perhaps mainstream secular science has learned something about the workings of God that God failed to reveal to any of his prophets? – choosing rather to reveal them to Darwin (who died as an agnostic without hope in a personal God because of his belief in naturalistic evolution as the source of the diversity of life)? What does the evidence that is actually available to us suggest? – that Darwin was right after all? Perhaps the church is also being inconsistent with science by trying to hold onto a literal interpretation of the story of Genesis with regard to the origin of life on this planet, but not so much with regard to the origin of the universe?
Beyond this, however, how good are the arguments presented by Dr. Rogers? Perhaps the church is wrong after all? Perhaps I should leave the SDA Church and Christianity as well? Perhaps mainstream secular science has learned something about the workings of God that God failed to reveal to any of his prophets? – choosing rather to reveal them to Darwin (who died as an agnostic without hope in a personal God because of his belief in naturalistic evolution as the source of the diversity of life)? What does the evidence that is actually available to us suggest? – that Darwin was right after all? Perhaps the church is also being inconsistent with science by trying to hold onto a literal interpretation of the story of Genesis with regard to the origin of life on this planet, but not so much with regard to the origin of the universe?
 First off, let’s consider the overall theological implications of life existing and evolving on this planet for a couple billion years or so. Rogers argues that its no big deal as long as one accepts all the waste associated with an ancient evolving universe. Well, to start with there’s a big difference between a rock and a sentient living thing. We’re not talking about bacteria or orange peals here. The key point to consider is that a rock can’t consciously experience pain or suffering or fear or death. That’s a big deal because a God who deliberately created using a protracted and very selfish and painful process of “survival of the fittest” that He knew would cause untold suffering and death for untold billions upon billions of sentient creatures would be seen, by most rational people anyway, as the very definition of evil. Such a notion also undermines the very concept of a “Moral Fall” and rebellion of mankind and the need for Jesus, the Son of God and Creator, to Himself come and selflessly die on the cross as the Savior of mankind – when He was the One who originally designed that all the suffering and pain and evil of this world should exist to begin with. This whole concept undermines the very fabric and logic of Christianity at large and paints God in the worst possible light.
First off, let’s consider the overall theological implications of life existing and evolving on this planet for a couple billion years or so. Rogers argues that its no big deal as long as one accepts all the waste associated with an ancient evolving universe. Well, to start with there’s a big difference between a rock and a sentient living thing. We’re not talking about bacteria or orange peals here. The key point to consider is that a rock can’t consciously experience pain or suffering or fear or death. That’s a big deal because a God who deliberately created using a protracted and very selfish and painful process of “survival of the fittest” that He knew would cause untold suffering and death for untold billions upon billions of sentient creatures would be seen, by most rational people anyway, as the very definition of evil. Such a notion also undermines the very concept of a “Moral Fall” and rebellion of mankind and the need for Jesus, the Son of God and Creator, to Himself come and selflessly die on the cross as the Savior of mankind – when He was the One who originally designed that all the suffering and pain and evil of this world should exist to begin with. This whole concept undermines the very fabric and logic of Christianity at large and paints God in the worst possible light.
But, perhaps God really is evil? Perhaps science has detected the work of an evil genius at play in our universe? If that is the case, I for one would like to know. After all, isn’t that what the “Great Controversy” is supposed to be all about? – putting God on trial before the universe to judge if He is really the God of love He claims to be? So, let’s look at some of the “science” cited by Dr. Rogers and see if God really is as evil as Rogers makes Him out to be…
 Biological Evolution: Dr. Rogers brings up the old argument that eukaryotic organisms (which includes humans) originally evolved hundreds of millions of years ago through a symbiotic relationship with bacteria. These internalized bacteria in turn eventually evolved into mitochondria – which are currently the source of the energy currency (in the form of the ATP molecule) of the eukaryotic cell.
Biological Evolution: Dr. Rogers brings up the old argument that eukaryotic organisms (which includes humans) originally evolved hundreds of millions of years ago through a symbiotic relationship with bacteria. These internalized bacteria in turn eventually evolved into mitochondria – which are currently the source of the energy currency (in the form of the ATP molecule) of the eukaryotic cell.
Now, it is true that mitochondrial organelles are quite unique and very interesting. Unlike any other organelle, except for chloroplasts, mitochondria appear to originate only from other mitochondria. They contain some of their own DNA, which is usually, but not always, circular – like circular bacterial DNA (there are also many organisms that have linear mitochondrial chromosomes with eukaryotic-style telomeres). Mitochondria also have their own transcriptional and translational machinery to decode DNA and messenger RNA and produce proteins. Also, mitochondrial ribosomes and transfer RNA molecules are similar to those found in bacteria,  as are some of the components of their membranes. In 1970, these and other similar observations led Dr. Lynn Margulis to propose an extracellular origin for mitochondria in her book, Origin of Eukaryotic Cells (Margulis, 1970). However, despite having their own DNA, mitochondria do not contain anywhere near the amount of DNA needed to code for all mitochondria-specific proteins. Over 99% of the proteins needed for mitochondrial function are actually produced outside of the mitochondria themselves. The DNA needed to code for these proteins is located within the cell’s nucleus and the protein sequences are assembled in the cytoplasm of the cell before being imported into the mitochondria (Endo and Yamano, 2010). It is hypothesized that these necessary genes were once part of the mitochondrial genome, but were then transferred and incorporated into the eukaryotic nuclear DNA over time. Not surprisingly then, none of the initial mtDNAs investigated by detailed sequencing, including animal mtDNAs, look anything like a typical bacterial genome in the way in which genes are organized and expressed (Michael Gray, 2012).
as are some of the components of their membranes. In 1970, these and other similar observations led Dr. Lynn Margulis to propose an extracellular origin for mitochondria in her book, Origin of Eukaryotic Cells (Margulis, 1970). However, despite having their own DNA, mitochondria do not contain anywhere near the amount of DNA needed to code for all mitochondria-specific proteins. Over 99% of the proteins needed for mitochondrial function are actually produced outside of the mitochondria themselves. The DNA needed to code for these proteins is located within the cell’s nucleus and the protein sequences are assembled in the cytoplasm of the cell before being imported into the mitochondria (Endo and Yamano, 2010). It is hypothesized that these necessary genes were once part of the mitochondrial genome, but were then transferred and incorporated into the eukaryotic nuclear DNA over time. Not surprisingly then, none of the initial mtDNAs investigated by detailed sequencing, including animal mtDNAs, look anything like a typical bacterial genome in the way in which genes are organized and expressed (Michael Gray, 2012).
It is interesting to note at this point that Margulis herself wasn’t really very Darwinian in her thinking. She opposed competition-oriented views of evolution and stressed the importance of symbiotic or cooperative relationships between species. She also argued that standard neo-Darwinism, which insists on the slow accrual of mutations by gene-level natural selection, “is in a complete funk” (Link).
But what about all of those similarities between mitochondria and bacteria? It would seem like these similarities should overwhelmingly support the theory of common ancestry between bacteria and mitochondria.
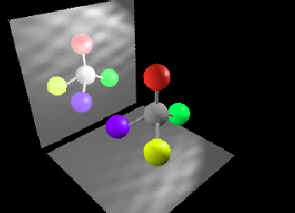
 Well, the problem with Darwinian thinking in general is that too much emphasis is placed on the shared similarities between various creatures without sufficient consideration of the uniquely required functional differences. These required differences are what the Darwinian mechanism cannot reasonably explain beyond the lowest levels of functional complexity (or minimum structural threshold requirements). The fact of the matter is that no one has ever observed nor has anyone ever published a reasonable explanation for how random mutations combined with natural selection can produce any qualitatively novel protein-based biological system that requires more than a few hundred specifically arranged amino acid residues – this side of trillions upon trillions of years of time. Functionally complex systems that require a minimum of multiple proteins comprised of several thousand specifically-coded amino acid residue positions, like a rotary flagellar motility system or ATPsynthase (illustrated), simply don’t evolve. It just doesn’t happen nor is it remotely likely to happen in what anyone would call a reasonable amount of time (Link). And, when it comes to mitochondria, there are various uniquely functional features that are required for successful symbiosis – that bacteria simply do not have. In other words, getting a viable symbiotic relationship established to begin with isn’t so simple from a purely naturalistic perspective.
Well, the problem with Darwinian thinking in general is that too much emphasis is placed on the shared similarities between various creatures without sufficient consideration of the uniquely required functional differences. These required differences are what the Darwinian mechanism cannot reasonably explain beyond the lowest levels of functional complexity (or minimum structural threshold requirements). The fact of the matter is that no one has ever observed nor has anyone ever published a reasonable explanation for how random mutations combined with natural selection can produce any qualitatively novel protein-based biological system that requires more than a few hundred specifically arranged amino acid residues – this side of trillions upon trillions of years of time. Functionally complex systems that require a minimum of multiple proteins comprised of several thousand specifically-coded amino acid residue positions, like a rotary flagellar motility system or ATPsynthase (illustrated), simply don’t evolve. It just doesn’t happen nor is it remotely likely to happen in what anyone would call a reasonable amount of time (Link). And, when it comes to mitochondria, there are various uniquely functional features that are required for successful symbiosis – that bacteria simply do not have. In other words, getting a viable symbiotic relationship established to begin with isn’t so simple from a purely naturalistic perspective.
 Consider, for example, that bacteria are not in the habit of sharing the ATP molecules they produce. Their ATPsynthase machine is set up so that the precious ATP molecules that are produced are released on the inside of the bacterium and the bacterium does not export the ATP that it makes. Of course, this same setup is also found in the mitochondria, but with one key difference. Mitochondria do export ATP for use by the host cell – but how? Now, both ATP and ADP (from which ATP is made) are highly charged molecules that do not diffuse freely across the inner mitochondrial membrane. So, how do the ATP molecules get outside of the mitochonria once they are produced so that they can be used by the host cell? Well, this requires a very specialized protein translocator protein (ATP/ADP Translocase; also known as Adenine nucleotide translocator or ANT) that moves the ATP molecules from the inner “matrix” of the mitochondria outward to the “intermembrane space” in exchange for ADP molecules (which are moved in the opposite direction). This system is set up so that the entry of ADP into the matrix is precisely coupled to the exit of ATP. ATP/ADP exchange is energetically expensive; about a quarter of the energy yield from electron transfer by the respiratory chain is consumed to regenerate the membrane potential that is consumed during this exchange process (Berg et al., 2002). And, this ANT machine is comprised of a homodimer with each subunit consisting of 297 residues and weighing approximately 30 kDa. Now that ATP is in the intermembrane space, it has to pass through another hurdle or gate-keeper channel known as the “voltage-gated channel” or VDAC (mitochondrial porin) – which is also a fairly complex in both structure and function (Dyson and Forte, 2008). As it turns out, ATP translocation through VDAC is facilitated by a set of specific in
Consider, for example, that bacteria are not in the habit of sharing the ATP molecules they produce. Their ATPsynthase machine is set up so that the precious ATP molecules that are produced are released on the inside of the bacterium and the bacterium does not export the ATP that it makes. Of course, this same setup is also found in the mitochondria, but with one key difference. Mitochondria do export ATP for use by the host cell – but how? Now, both ATP and ADP (from which ATP is made) are highly charged molecules that do not diffuse freely across the inner mitochondrial membrane. So, how do the ATP molecules get outside of the mitochonria once they are produced so that they can be used by the host cell? Well, this requires a very specialized protein translocator protein (ATP/ADP Translocase; also known as Adenine nucleotide translocator or ANT) that moves the ATP molecules from the inner “matrix” of the mitochondria outward to the “intermembrane space” in exchange for ADP molecules (which are moved in the opposite direction). This system is set up so that the entry of ADP into the matrix is precisely coupled to the exit of ATP. ATP/ADP exchange is energetically expensive; about a quarter of the energy yield from electron transfer by the respiratory chain is consumed to regenerate the membrane potential that is consumed during this exchange process (Berg et al., 2002). And, this ANT machine is comprised of a homodimer with each subunit consisting of 297 residues and weighing approximately 30 kDa. Now that ATP is in the intermembrane space, it has to pass through another hurdle or gate-keeper channel known as the “voltage-gated channel” or VDAC (mitochondrial porin) – which is also a fairly complex in both structure and function (Dyson and Forte, 2008). As it turns out, ATP translocation through VDAC is facilitated by a set of specific in
teractions between ATP and the channel wall (Komarov, 2005).
 The problem, of course, is that it is hard to imagine much of an endosymbiotic advantage for a primitive eukaryotic cell engulfing a bacterium and keeping it around for a while if the bacterium wasn’t providing an immediate advantage. After all, if the bacterium isn’t sharing its own ATP, and it is using the host’s resources to generate that ATP, the relationship between them would seem to be more parasitic than symbiotic. And, as it turns out, the complex and highly specific ANT translocation machine is not found in any bacterial genome with the exception of parasitic Rickettsia and Chlamydia bacteria (Stephan Schmitz-Esser, 2004).
The problem, of course, is that it is hard to imagine much of an endosymbiotic advantage for a primitive eukaryotic cell engulfing a bacterium and keeping it around for a while if the bacterium wasn’t providing an immediate advantage. After all, if the bacterium isn’t sharing its own ATP, and it is using the host’s resources to generate that ATP, the relationship between them would seem to be more parasitic than symbiotic. And, as it turns out, the complex and highly specific ANT translocation machine is not found in any bacterial genome with the exception of parasitic Rickettsia and Chlamydia bacteria (Stephan Schmitz-Esser, 2004).
So, the big question is, where did the Rickettsia and Chlamydia bacteria get these genes for the ANT translocation system? – since no other bacteria have this system and these particular bacteria have lost most of the genes that other free-living bacteria have? Well, as it turns out, they got the specialized ANT genes by horizontal transfer from plants. In other words, they are “plastid homologues” unrelated to mitochondrial ANT genes (Kurland and Andersson, 2000). In fact, there are several other cases of acquisition of eukaryotic genes by bacteria – to include isoleucyl-tRNA synthetases whose acquisition from eukaryotes by several bacteria is linked to antibiotic resistance, and proteases that may be implicated in the pathogenesis of Chlamydia. (Koonin et al. 2001). In fact, the acquisition of eukaryotic genes by bacteria is a well-known phenomenon – especially parasites (Link).
 So, how do the ANT-like genes (TLC genes) turn the Rickettsia and Chlamydia bacteria into parasites? – when mitochondria are not parasitic? Well, the Rickettsia and Chlamydia bacteria turn the process on its head so that instead of using the system to export ATP, they use it to import ATP from the host in exchange for exporting ADP – thereby sapping the host cell of energy.
So, how do the ANT-like genes (TLC genes) turn the Rickettsia and Chlamydia bacteria into parasites? – when mitochondria are not parasitic? Well, the Rickettsia and Chlamydia bacteria turn the process on its head so that instead of using the system to export ATP, they use it to import ATP from the host in exchange for exporting ADP – thereby sapping the host cell of energy.
In short, the Rickettsial-type system (from which mitochondria supposedly evolved) originally gained the necessary genes, not from previous evolution or from some other type of bacterial species, but from eukaryotic plants! In other words, eukaryotic organisms already had the necessary genes to begin with, which were then stolen by bacteria, allowing them to become parasitic. And, VDAC-type porins are also found in bacteria (Godbole, 2011), but, like ANT genes in bacteria, are often used for pathogenic purposes (Anne Müller, 2002).
 This is a form of degenerative evolution. It’s not a form of enhanced symbiotic functional complexity at all. In fact, there are many other examples of systems within parasitic organisms that originally arose, not by the evolution of simpler systems, but by the devolution or loss of informational complexity of more complex systems. Consider the Type III Secretory System (TTSS) used by pathologic bacteria as a toxin injector (bacteria such as Yersinia pestis, which are responsible for bubonic plague). For many years it was argued by prominent evolutionists (like Kenneth Miller or Nick Matzke) that the very functionally complex rotary bacterial flagellar motility system evolved from the TTSS system or something like it. It is now known, however, that the TTSS system devolved from the fully formed and functional flagellar motility system – not the other way around (Toft and Fares, 2008).
This is a form of degenerative evolution. It’s not a form of enhanced symbiotic functional complexity at all. In fact, there are many other examples of systems within parasitic organisms that originally arose, not by the evolution of simpler systems, but by the devolution or loss of informational complexity of more complex systems. Consider the Type III Secretory System (TTSS) used by pathologic bacteria as a toxin injector (bacteria such as Yersinia pestis, which are responsible for bubonic plague). For many years it was argued by prominent evolutionists (like Kenneth Miller or Nick Matzke) that the very functionally complex rotary bacterial flagellar motility system evolved from the TTSS system or something like it. It is now known, however, that the TTSS system devolved from the fully formed and functional flagellar motility system – not the other way around (Toft and Fares, 2008).  Consider this in the light of the fact that almost all of the proteins used to build the rotary bacterial flagellum are homologous to one degree or another to other bacterial proteins that are doing other jobs as parts of other systems of function. However, homology alone doesn’t explain how all the modifications that are necessary to get all of these various proteins together to produce an entirely new and unique collective function. Until all of these required modifications are achieved, the next proposed beneficial steppingstone in the evolutionary pathway simply cannot be realized to any functional advantage that nature can select for in a positive manner (Link).
Consider this in the light of the fact that almost all of the proteins used to build the rotary bacterial flagellum are homologous to one degree or another to other bacterial proteins that are doing other jobs as parts of other systems of function. However, homology alone doesn’t explain how all the modifications that are necessary to get all of these various proteins together to produce an entirely new and unique collective function. Until all of these required modifications are achieved, the next proposed beneficial steppingstone in the evolutionary pathway simply cannot be realized to any functional advantage that nature can select for in a positive manner (Link).
The same problem is true for the mitochondrial system. Homologies don’t explain the necessary functional differences. Consider, for example, that around 42% of the proteins participating in the aerobic ATP-generating system of yeast mitochondria have been found only in eukaryotes (Kurland and Andersson, 2000). Also, “the protein translocases that drive protein import into mitochondria have no obvious counterparts in bacteria” (Dolzal, et al., 2006). And, these numerous required functional differences at these higher levels of functional complexity (I’ve only highlighted a couple of them here) are what help one detect the required signature of a very intelligent Designer behind these functionally complex systems.
How so? Because, beyond the lowest levels of functional complexity, biological systems, like any other functionally complex machine or functional computer code where multiple specific orientations of building blocks or code are required, simply don’t evolve and are very unlikely to have ever evolved on this planet or anywhere else in the universe since the universe. There’s just not remotely enough time by anyone’s book to make the statistical odds tenably favor the Darwinian mechanism when it comes to these higher level systems. What’s left is just a story about how the evolution of this relationship must have happened. However, there’s really no science, no viable mechanism much less any kind of meaningful demonstration, to back it up. It’s all based on the assumption that similarities between various living things must mean common evolutionary ancestry – not common design.
 Theory of Plate tectonics:
Theory of Plate tectonics:
While the movement of the continental plates does seem quite obvious and well beyond serious debate (i.e., the continents do seem to fit together like puzzle pieces), what is not so clear is Dr. Roger’s assertion that this process required vast periods of time. While it is true that continental drift is currently taking place at a rather sedate pace, if 200 million years had really gone by since the continents were last connected, where is all the continental erosion, from above and from all sides, that should have taken place during that time? The pieces of the puzzle should no longer fit.
Also where did all the expected sediment go that should be present within the ocean basins if they are really that old? The math is fairly straightforward here: We know how much sediment is being deposited in the oceans on a yearly basis (~30 billion metric tons). We know how much sediment is being removed from the oceans by subduction (~2 billion tons). This leaves ~28 billion tons to increase on the ocean floor every year. Right off the bat this presents a problem for the non-catastrophic models of plate tectonics. After all, at the rate of 28 billion tons per year (~13 km3 of sediment) all of the ocean basins (total volume of ~1550 million km3) could be completely filled with sediment in just under 120 million years. It seems, rather, that continental drift was much more rapid in the past than it is today – as would be expected from a sudden large catastrophic release of energy (which would also be consistent with a worldwide watery catastrophe of Noachian proportions).
Also, recent evidence suggest far more rapid magnetic reversals in the past than can be explained by non-catastrophic models of plate tectonics (Link).
For further discussion see: Link, Link
 Geologic Column:
Geologic Column:
The geologic column certainly is interesting and I’m not saying that all of its features are without any mystery from a creationist perspective. However, I dare say that it presents old age uniformitarians with far greater and more significant mysteries in comparison. One major difficulty is the general lack of erosion between the layers that would be expected if these sedimentary layers of rock were originally laid down over long periods of time with vast stretches of time transpiring between the layers. There is also the problem of a general lack of the expected degree of bioturbation (sediment mixing by burrowing organisms), worldwide, within the geologic column. Continental or even worldwide paleocurrents all pointing in the same general direction for a given series of layers (Link) are also much easier for a catastrophic Flood model to explain. And, creationist models have proven themselves to have good predictive value when it comes to debunking various mainstream interpretations of various formations within the geologic record. For example, Dr. Ariel Roth demonstrated that what were once thought to be “termite nests” within the layers of the Jurassic Morrison Formation (pretty hard to explain with a Flood model) were actually inorganic formations – not termite nests at all (Link). And, there are many more such problems that seem to strongly favor or are at least more consistent with a catastrophic Flood model.
So, while “hydrological sorting, ecological zonation, and relative mobility” may not explain all the mysteries of the geologic column or fossil records, they do explain some features quite well and other factors are being discovered that appear to explain more and more mysteries from a Biblical young-life perspective.
For more information see: Link, Link

Radiometric Dating:
While radiometric dating methods do seem to dramatically counter the claims of young-life creationists, it’s not all peaches and cream for those on the opposite side of the fence. Dr. Rogers doesn’t seem to recognize the fact that radiocarbon dating, in particular, strongly favors the young-life position since significant quantities of radiocarbon have been discovered in the soft tissues of all dinosaur bones examined thus far, producing ages ranging from 16,000 to 32,000 years – essentially the same as the radiocarbon ages reported for large Pleistocene mammals such as mammoths, mastodons, dire wolves, etc. (Link, Link). Also, pretty much all coal samples contain fairly significant quantities of radiocarbon. The usual counterarguments of either contamination or in situ production don’t hold water when it comes to explaining the very high levels of radiocarbon so consistently found within the fossil record (Link).
 Also, Dr. Rogers doesn’t seem to be up to date with regard to amino acid racemization dating (AAR) – not realizing that it is no longer considered to be the independent dating technique that its father, Peter Edward Hare, originally believed it to be. It is now known to be a relative dating technique that must first be calibrated against radiocarbon dating for a given area of interest before it can be used. Also, since the racemization rate of amino acids is heavily temperature dependent, the thermal history of a given location must also be known (Link). Ironically, it ends up that the AAR dating technique is more in line with the young-life position as well and does not support the ancient arrival of life on this planet at all (Link).
Also, Dr. Rogers doesn’t seem to be up to date with regard to amino acid racemization dating (AAR) – not realizing that it is no longer considered to be the independent dating technique that its father, Peter Edward Hare, originally believed it to be. It is now known to be a relative dating technique that must first be calibrated against radiocarbon dating for a given area of interest before it can be used. Also, since the racemization rate of amino acids is heavily temperature dependent, the thermal history of a given location must also be known (Link). Ironically, it ends up that the AAR dating technique is more in line with the young-life position as well and does not support the ancient arrival of life on this planet at all (Link).
In short, there are many problems with the mainstream interpretations of various radiometric dating methods and, while some mysteries do still remain, radiometric dating is not at all the nail in the coffin for the young-life position that Rogers makes it out to be (Link).
 Ice Core Dating:
Ice Core Dating:
The idea of ice core dating seems simple and straightforward enough – very much like tree-ring dating or the dating of sedimentary layers or varves within lake beds. All you have to do is count the layers and that will give you the correct age. But, it’s not that simple. With ice core dating in particular, the layers get compressed and stretched and thinned until they simply cannot be recognized with the naked eye. At this point, the layers are detected by chemical analysis or annual signatures. The problem with these chemical signatures is that the chemicals themselves do not say put in the ice layers. They move and form pseudo-layers (Link). Beyond this, multiple layers can be deposited in a single year, which complicates the issue.
Radiocarbon dating helps a bit, but it ends up being that the ice core layers themselves are not very helpful as a source of independent age calibration. The same is true of other layer-based age calculations. For example, lake varves, along with tree rings, etc., have long been used to “calibrate” C14 dates (Link). The reason for this need for calibration is because the ratio of C14/C12 in the atmosphere doesn’t stay the same. It changes and has changed significantly over time (Link). That means, of course, that if you date something by C14 methods that is also used to calibrate the C14 dating method, you’re obviously going to get a “straight line” agreement. It’s a circular argument. And, as with counting tree rings and ice core layers, varve layers are not reliable annual markers to use as an independent means of calibrating C14.
 It is also interesting to note that Greenland, in particular, has not always been covered in ice. It was once truly green – all over. In fact, within the Hypsithermal period or “warm age” (which is said to have lasted some 7,000 years, ending only some just 2,500 years ago), the northernmost parts of the planet were very much warmer than they are today. Studies on sedimentary cores carried out in the North Atlantic between Hudson Strait and Cape Hatteras indicate ocean temperatures of 18°C (verses about 8°C today in this region) during the height of this period of time between 4,000 to 6,000 years ago in mainstream thinking. Given that the Greenland ice sheet is currently melting at a fairly rapid rate, it’s rather hard to believe that it existed at all during the very warm Hypsithermal period – a periods when millions of mammoths along with many other types of warmer weather plants and animals happily lived within the Arctic Circle all around the globe along the very same latitudes as Greenland (Link). Carbon 14 dating of Siberian mammoths has shown that they lived in this region till about 9,670 years ago. A 1995 study of mammoth remains located on Wrangel Island (on the border of the East-Siberian and Chukchi Seas) shows that mammoths persisted on this island till about 1,700 B.C. (Vartanyan S.L, et. al., 1995). And yet, somehow, Greenland was still covered with thick sheets of ice when everything around it was warm and balmy, supporting huge herds of animals and lush forests with fruit bearing trees and abundant grasslands?
It is also interesting to note that Greenland, in particular, has not always been covered in ice. It was once truly green – all over. In fact, within the Hypsithermal period or “warm age” (which is said to have lasted some 7,000 years, ending only some just 2,500 years ago), the northernmost parts of the planet were very much warmer than they are today. Studies on sedimentary cores carried out in the North Atlantic between Hudson Strait and Cape Hatteras indicate ocean temperatures of 18°C (verses about 8°C today in this region) during the height of this period of time between 4,000 to 6,000 years ago in mainstream thinking. Given that the Greenland ice sheet is currently melting at a fairly rapid rate, it’s rather hard to believe that it existed at all during the very warm Hypsithermal period – a periods when millions of mammoths along with many other types of warmer weather plants and animals happily lived within the Arctic Circle all around the globe along the very same latitudes as Greenland (Link). Carbon 14 dating of Siberian mammoths has shown that they lived in this region till about 9,670 years ago. A 1995 study of mammoth remains located on Wrangel Island (on the border of the East-Siberian and Chukchi Seas) shows that mammoths persisted on this island till about 1,700 B.C. (Vartanyan S.L, et. al., 1995). And yet, somehow, Greenland was still covered with thick sheets of ice when everything around it was warm and balmy, supporting huge herds of animals and lush forests with fruit bearing trees and abundant grasslands?
More at: Ancient Ice
And the list goes on and on. Not only is biological evolution via the mechanism of random mutations and natural selection simply untenable beyond the very lowest levels of functional complexity this side of trillions upon trillions of years of time (Link), the notion that life has existed, evolved, suffered, and died on this planet for hundreds of millions of years simply isn’t consistent with a great deal of very good scientific evidence – some of which, ironically, Rogers unknowingly cited himself.
But what about the charge of inconsistency? Is the church really being inconsistent with regard to its use of the story of Genesis and science at large? – in trying to interpret the origin of life on this planet as young while allowing for the ancient origin of the universe?
 Rogers suggests that the Adventist church started out teaching that the entire universe was created during the “creation week” described in Genesis – but this is hardly the case. The concept that there is a “gap in time” between Genesis 1:1 and Genesis 1:2 is known as the “Passive Gap Theory” – which was supported by many of the founding fathers of the SDA Church – including Ellen White. As a matter of fact, Mrs. White never says that the entire universe was created during our creation week. To the contrary, she specifically says that the universe pre-existed our creation week – and that angels and intelligences on other inhabited worlds witnessed the creation week of our planet (Job 38:7). This is consistent with the Passive Gap Theory where the “stars”, mentioned on Day 4 of our creation week, already existed and simply became visible through the atmosphere on Day 4 (as did the Sun and the moon).
Rogers suggests that the Adventist church started out teaching that the entire universe was created during the “creation week” described in Genesis – but this is hardly the case. The concept that there is a “gap in time” between Genesis 1:1 and Genesis 1:2 is known as the “Passive Gap Theory” – which was supported by many of the founding fathers of the SDA Church – including Ellen White. As a matter of fact, Mrs. White never says that the entire universe was created during our creation week. To the contrary, she specifically says that the universe pre-existed our creation week – and that angels and intelligences on other inhabited worlds witnessed the creation week of our planet (Job 38:7). This is consistent with the Passive Gap Theory where the “stars”, mentioned on Day 4 of our creation week, already existed and simply became visible through the atmosphere on Day 4 (as did the Sun and the moon).
In fact, this view was predominant among other Adventist pioneers as well. For example, M. C. Wilcox wrote in 1898:
“When did God create, or bring into existence, the heavens and the earth? ‘In the beginning.’ When this ‘beginning’ was, how long a period it covered, it is idle to conjecture; for it is not revealed. That it was a period which antedated the six days’ work is evident.”
M.C. Wilcox, “The Gospel in Genesis One,” The Signs of the Times, 24.27 (July 7, 1898): 16.
In fact, from the 1860s on, many of the pioneers of the Seventh-day Adventist Church, aside from J.N. Andrews, had been discussing this issue in favor of the Passive Gap Theory. Uriah Smith, editor of the Review and Herald, published several pages from a book or pamphlet in 1860, entitled The Bible True, as follows:
“Nor is there anything in revelation which forbids us to believe that the substance of the earth was formed long before it received its present organization. The first verse of Genesis may relate to a period millions of ages prior to the event noticed in the rest of the chapter. Commentators who wrote hundreds, and some of them fifteen hundred years ago, seem to have understood the first verse as relating to a period far anterior to the creation of man. This interpretation, therefore, is not modern, nor made merely to obviate a difficulty. But if it were, it is so perfectly coincident with the just rules of interpretation, that there can be no just objection to it.” – Uriah Smith

Consider also the more recent comments of Richard Davidson along these lines:
Against the suggestion that all the words in Genesis 1:2 simply imply “nothingness,” it must be observed that verses 3 and following do not describe the creation of water, but assume its prior existence. The word tehom “deep,” combined with tohu and bohu together (as in Jeremiah 4:34) do not seem to refer to nothingness, but rather to the earth in an unformed-unfilled state covered by water.
This leads us to the initial unformed-unfilled position. A straightforward reading of the flow of thought in Genesis 1:1-3 has led the majority of Christian and Jewish interpreters in the history of interpretation to this position, hence this is called the traditional view.
Richard Davidson (Dialogue)

So, the charge of inconsistency with regard to the literal understanding of the story of Genesis or even the weight of scientific evidence doesn’t hold water. Rather, it seems like Rogers is the one who is trying to have his cake and eat it too. He’s trying to maintain some basis for belief in God and at least some value or usefulness for the Bible itself while, at the same time, trying to incorporate mainstream Darwinian thinking. The fact is that these two views are diametrically opposed. Both cannot rationally be true. If the Darwinian story of origins is true, then the claims of the Bible, regarding all that Christianity has to offer, are simply ridiculous – completely non-credible.
For example, William Provine, late professor of biological sciences at Cornell University, gave a very interesting speech for a 1998 Darwin Day keynote address in which he pointed out the following:
Naturalistic evolution has clear consequences that Charles Darwin understood perfectly.
- No gods worth having exist;
- No life after death exists;
- No ultimate foundation for ethics exists;
- No ultimate meaning in life exists; and
- Human free will is nonexistent.
Provine, William B. [Professor of Biological Sciences, Cornell University], “Evolution: Free will and punishment and meaning in life”, Abstract of Will Provine’s 1998 Darwin Day Keynote Address.

Provine also wrote, “In other words, religion is compatible with modern evolutionary biology (and indeed all of modern science) if the religion is effectively indistinguishable from atheism.” – Academe January 1987, pp.51-52
It seems to me that Provine, contrary to the claim forwarded by Rogers, was right and was most consistent with the implications of accepting neo-Darwinian claims. Darwinian-style evolution is just one more argument for the philosophical position of “Naturalism” – a position that suggests that everything within the physical world, everything that we can see, touch, hear, taste, or smell, is ultimately the result of non-deliberate mindless forces of nature. These forces do not have feelings or care about you or me or our feelings regarding what they are or are not doing to us or for us.
Beyond this, however, I’m not sure why the leadership of the Seventh-day Adventist Church doesn’t say and/or do more about the teachings of the schools that bear its name? At the very least, if the direction of one of our church schools cannot be put back in line with the primary goals and ideas of the church organization, why continue to lend the church’s name to such a school?





